



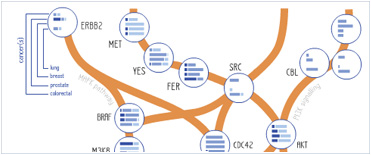







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
Phospho-Ser9 Synapsin I Antibody
Affinity purified rabbit polyclonal antibody

- SPECIFICATION
- CITATIONS: 1
- PROTOCOLS
- BACKGROUND

Application
| WB, IHC |
|---|---|
| Primary Accession | P17599 |
| Reactivity | Rat |
| Predicted | Bovine, Human, Mouse, Xenopus, Zebrafish |
| Host | Rabbit |
| Clonality | polyclonal |
| Calculated MW | 78 KDa |
| Gene ID | 281510 |
|---|---|
| Gene Name | SYN1 |
| Other Names | Synapsin-1, Synapsin I, SYN1 |
| Target/Specificity | Synthetic phospho-peptide corresponding to amino acid residues surrounding Ser9 conjugated to KLH. |
| Dilution | WB~~ 1:1000 IHC~~ 1:500 |
| Format | Prepared from rabbit serum by affinity purification via sequential chromatography on phospho- and dephosphopeptide affinity columns. |
| Antibody Specificity | Specific for ~78k synapsin I doublet protein phosphorylated at Ser9. Theantibody also weakly labels the ~55k synapsin II protein which has a similar phosphorylationsite to that of Ser9on synapsin I. Immunolabeling is blocked by preadsorption of the antibodywith the phosphopeptide used as antigen but not by the corresponding dephosphopeptide.Immunolabeling is also completely eliminated by treatment withλphosphatase. |
| Storage | Maintain refrigerated at 2-8°C for up to 6 months. For long term storage store at -20°C in small aliquots to prevent freeze-thaw cycles. |
| Precautions | Phospho-Ser9 Synapsin I Antibody is for research use only and not for use in diagnostic or therapeutic procedures. |
| Shipping | Blue Ice |

Provided below are standard protocols that you may find useful for product applications.
Background
Synapsin I plays a key role in synaptic plasticity in brain (Feng et al., 2002; Nayak et al., 1996). This effect is due in large part to the ability of the synapsins to regulate the availability of synaptic vesicles for release. In addition to its role in plasti city, the expression of synapsin I is a precise indicator of synapse formation (Moore and Bernstein, 1989; Stone et al., 1994). Thus, synapsin I immunocytochemistry provides a valuable tool for the study of synaptogenesis. The role of synapsin in synaptic plasticity and in synaptogensis is regulated by phosphor ylation (Jovanovic et al., 2001; Kao et al., 2002). Serine 9 is the site on synapsin I that is phosphorylated by cAMP-dependent protein kinase and by calcium calmodulin kinase I (Czernik et al., 1987). Phosphorylation of this site is thought to regulate synaptic vesicle function and neurite outgrowth (Kao et al., 2002).
References
Czernik AJ, Pang DT, Greengard P (1987) Amino acid
sequences surrounding the cAMP-dependent and
calcium/calmodulin-dependent phosphorylation sites in rat and bovine synapsin I. Proc Natl Acad Sci (USA)
84:7518-7522.
Feng J, Chi P, Blanpied TA, Xu YM, Magarinos AM, Fe
rreira A, Takahashi RH, Kao HT, McEwen BS, Ryan TA,
Augustine GJ, Greengard P (2002) Regulation of neurotransmitter release by synapsin III. J Neurosci 22:4372-
4380.
Jovanovic JN, Sihra TS, Nairn AC, Hemmings HC, Jr., Gr
eengard P, Czernik AJ (2001) Opposing changes in
phosphorylation of specific sites in synapsin I during Ca
2+
-dependent glutamate release in isolated nerve
terminals. J Neurosci 21:7944-7953.
Kao HT, Song HJ, Porton B, Ming GL, Hoh J, Abraham M,
Czernik AJ, Pieribone VA, Poo MM, Greengard P (2002) A
protein kinase A-dependent molecular switch in synapsin
s regulates neurite outgrowth. Nature Neurosci 5:431-
437.
Moore RY, Bernstein M (1989) Synaptogenesis in the rat suprachiasmatic nucleus demonstrated by electron
microscopy and synapsin I immunoreactivity. J Neurosci 9:2151-2162.
Nayak AS, Moore CI, Browning MD (1996) CaM Kinase II
phosphorylation of the presyn
aptic protein synapsin is
persistently increased during expression of long-term po
tentiation. Proc Natl Acad Sci (USA) 93:15451-15456.
Stone LM, Browning MD, Finger TE (1994) Differential dist
ribution of the synapsins in the rat olfactory bulb. J
Neurosci 14:301-309.
Sachiko Shimomura, Tadashi Nagamine, Naoya Hatano, Noriyuki Sueyoshi, and Isamu Kameshita (2010)
Identification of an endogenou
s substrate of zebrafish doublecortin-like
protein kinase using a highly active
truncation mutant
.
J. Biochem.,
147: 711 - 722.
Note: Dr. Michael Browning co-author of t
he cited papers is the President and founder of
PhosphoSolutions.
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Other Products
Shipping Information
