



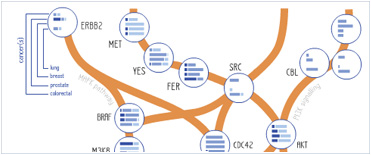







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
SUV39H1 Antibody
Purified Rabbit Polyclonal Antibody (Pab)

- SPECIFICATION
- CITATIONS: 2
- PROTOCOLS
- BACKGROUND

Application
| WB, E |
|---|---|
| Primary Accession | O43463 |
| Other Accession | NP_003164 |
| Reactivity | Human, Mouse |
| Host | Rabbit |
| Clonality | Polyclonal |
| Isotype | Rabbit IgG |
| Calculated MW | 47907 Da |
| Gene ID | 6839 |
|---|---|
| Other Names | Histone-lysine N-methyltransferase SUV39H1, Histone H3-K9 methyltransferase 1, H3-K9-HMTase 1, Lysine N-methyltransferase 1A, Position-effect variegation 3-9 homolog, Suppressor of variegation 3-9 homolog 1, Su(var)3-9 homolog 1, SUV39H1, KMT1A, SUV39H |
| Target/Specificity | Purified recombinant GST fusion protein encoding N-terminal of human SUV39H1. |
| Dilution | WB~~1:1000 |
| Format | Purified polyclonal antibody supplied in PBS with 0.09% (W/V) sodium azide. This antibody is prepared by Saturated Ammonium Sulfate (SAS) precipitation followed by dialysis against PBS. |
| Storage | Maintain refrigerated at 2-8°C for up to 2 weeks. For long term storage store at -20°C in small aliquots to prevent freeze-thaw cycles. |
| Precautions | SUV39H1 Antibody is for research use only and not for use in diagnostic or therapeutic procedures. |
| Name | SUV39H1 |
|---|---|
| Synonyms | KMT1A, SUV39H |
| Function | Histone methyltransferase that specifically trimethylates 'Lys-9' of histone H3 using monomethylated H3 'Lys-9' as substrate. Also weakly methylates histone H1 (in vitro). H3 'Lys-9' trimethylation represents a specific tag for epigenetic transcriptional repression by recruiting HP1 (CBX1, CBX3 and/or CBX5) proteins to methylated histones. Mainly functions in heterochromatin regions, thereby playing a central role in the establishment of constitutive heterochromatin at pericentric and telomere regions. H3 'Lys-9' trimethylation is also required to direct DNA methylation at pericentric repeats. SUV39H1 is targeted to histone H3 via its interaction with RB1 and is involved in many processes, such as repression of MYOD1-stimulated differentiation, regulation of the control switch for exiting the cell cycle and entering differentiation, repression by the PML-RARA fusion protein, BMP-induced repression, repression of switch recombination to IgA and regulation of telomere length. Component of the eNoSC (energy-dependent nucleolar silencing) complex, a complex that mediates silencing of rDNA in response to intracellular energy status and acts by recruiting histone-modifying enzymes. The eNoSC complex is able to sense the energy status of cell: upon glucose starvation, elevation of NAD(+)/NADP(+) ratio activates SIRT1, leading to histone H3 deacetylation followed by dimethylation of H3 at 'Lys-9' (H3K9me2) by SUV39H1 and the formation of silent chromatin in the rDNA locus. Recruited by the large PER complex to the E-box elements of the circadian target genes such as PER2 itself or PER1, contributes to the conversion of local chromatin to a heterochromatin-like repressive state through H3 'Lys-9' trimethylation. |
| Cellular Location | Nucleus. Nucleus lamina. Nucleus, nucleoplasm Chromosome, centromere. Note=Associates with centromeric constitutive heterochromatin |

Provided below are standard protocols that you may find useful for product applications.
Background
Similar to acetylation and phosphorylation, histone methylation at the N-terminal tail has emerged as an important role in regulating chromatin dynamics and gene activity. Histone methylation occurs on arginine and lysine residues and is catalyzed by two families of proteins, the protein arginine methyltransferase family and the SET-domain-containing methyltransferase family. Five members have been identified in the arginine methyltransferase family. About 27 are grouped into the SET-domain family, and another 17 make up the PR domain family that is related to the SET domain family. The retinoblastoma protein-interacting zinc finger geneRIZ1 is a tumor suppressor gene and a FOUNDING member of the PR domain family. RIZ1 inactivation is commonly found in many types of human cancers and occurs through loss of mRNA expression, frame shift mutation, chromosomal deletion, and missense mutation. RIZ1 is also a tumor susceptibility gene in mice. The loss of RIZ1 mRNA in human cancers was shown to associate with DNA methylation of its promoter CpG island. Methylation of the RIZ1 promoter strongly correlated with lost or decreased RIZ1 mRNA expression in breast, liver, colon, and lung cancer cell lines as well as in liver cancer tissues.
References
Fujita, N., et al., J. Biol. Chem. 278(26):24132-24138 (2003). Macaluso, M., et al., Oncogene 22(23):3511-3517 (2003). Schotta, G., et al., EMBO J. 21(5):1121-1131 (2002). Vaute, O., et al., Nucleic Acids Res. 30(2):475-481 (2002). Rea, S., et al., Nature 406(6796):593-599 (2000).
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Other Products
Shipping Information
