


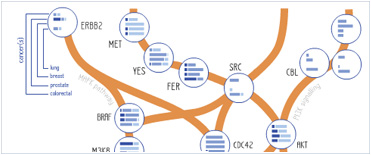







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
CUGBP1 Antibody (N-term)
Affinity Purified Rabbit Polyclonal Antibody (Pab)

- SPECIFICATION
- CITATIONS
- PROTOCOLS
- BACKGROUND

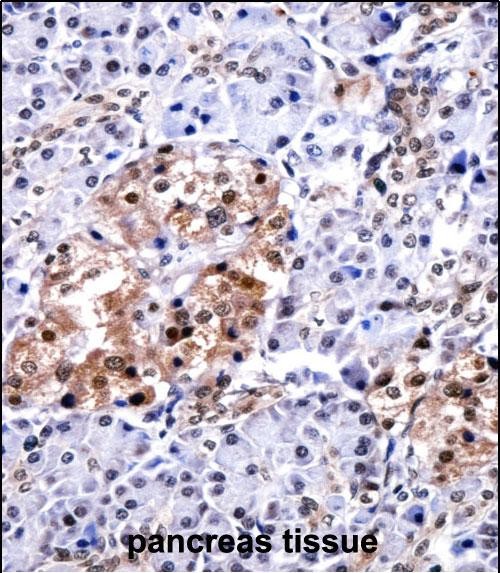
Application
| WB, IHC-P, E |
|---|---|
| Primary Accession | Q92879 |
| Other Accession | Q4QQT3, P28659, Q5F3T7, NP_001166110.1, NP_001020767.1, NP_941989.1, NP_001166111.1 |
| Reactivity | Human |
| Predicted | Chicken, Mouse, Rat |
| Host | Rabbit |
| Clonality | Polyclonal |
| Isotype | Rabbit IgG |
| Calculated MW | 52063 Da |
| Antigen Region | 1-30 aa |
| Gene ID | 10658 |
|---|---|
| Other Names | CUGBP Elav-like family member 1, CELF-1, 50 kDa nuclear polyadenylated RNA-binding protein, Bruno-like protein 2, CUG triplet repeat RNA-binding protein 1, CUG-BP1, CUG-BP- and ETR-3-like factor 1, Deadenylation factor CUG-BP, Embryo deadenylation element-binding protein homolog, EDEN-BP homolog, RNA-binding protein BRUNOL-2, CELF1, BRUNOL2, CUGBP, CUGBP1, NAB50 |
| Target/Specificity | This CUGBP1 antibody is generated from rabbits immunized with a KLH conjugated synthetic peptide between 1-30 amino acids from the N-terminal region of human CUGBP1. |
| Dilution | WB~~1:1000 IHC-P~~1:10~50 |
| Format | Purified polyclonal antibody supplied in PBS with 0.09% (W/V) sodium azide. This antibody is purified through a protein A column, followed by peptide affinity purification. |
| Storage | Maintain refrigerated at 2-8°C for up to 2 weeks. For long term storage store at -20°C in small aliquots to prevent freeze-thaw cycles. |
| Precautions | CUGBP1 Antibody (N-term) is for research use only and not for use in diagnostic or therapeutic procedures. |
| Name | CELF1 |
|---|---|
| Synonyms | BRUNOL2, CUGBP, CUGBP1, NAB50 |
| Function | RNA-binding protein implicated in the regulation of several post-transcriptional events. Involved in pre-mRNA alternative splicing, mRNA translation and stability. Mediates exon inclusion and/or exclusion in pre-mRNA that are subject to tissue-specific and developmentally regulated alternative splicing. Specifically activates exon 5 inclusion of cardiac isoforms of TNNT2 during heart remodeling at the juvenile to adult transition. Acts both as an activator and as a repressor of a pair of coregulated exons: promotes inclusion of the smooth muscle (SM) exon but exclusion of the non-muscle (NM) exon in actinin pre-mRNAs. Activates SM exon 5 inclusion by antagonizing the repressive effect of PTB. Promotes exclusion of exon 11 of the INSR pre-mRNA. Inhibits, together with HNRNPH1, insulin receptor (IR) pre- mRNA exon 11 inclusion in myoblast. Increases translation and controls the choice of translation initiation codon of CEBPB mRNA. Increases mRNA translation of CEBPB in aging liver (By similarity). Increases translation of CDKN1A mRNA by antagonizing the repressive effect of CALR3. Mediates rapid cytoplasmic mRNA deadenylation. Recruits the deadenylase PARN to the poly(A) tail of EDEN-containing mRNAs to promote their deadenylation. Required for completion of spermatogenesis (By similarity). Binds to (CUG)n triplet repeats in the 3'-UTR of transcripts such as DMPK and to Bruno response elements (BREs). Binds to muscle-specific splicing enhancer (MSE) intronic sites flanking the alternative exon 5 of TNNT2 pre-mRNA. Binds to AU-rich sequences (AREs or EDEN-like) localized in the 3'-UTR of JUN and FOS mRNAs. Binds to the IR RNA. Binds to the 5'-region of CDKN1A and CEBPB mRNAs. Binds with the 5'-region of CEBPB mRNA in aging liver. May be a specific regulator of miRNA biogenesis. Binds to primary microRNA pri-MIR140 and, with CELF2, negatively regulates the processing to mature miRNA (PubMed:28431233). |
| Cellular Location | Nucleus. Cytoplasm. Note=RNA-binding activity is detected in both nuclear and cytoplasmic compartments |
| Tissue Location | Ubiquitous. |

Thousands of laboratories across the world have published research that depended on the performance of antibodies from Abcepta to advance their research. Check out links to articles that cite our products in major peer-reviewed journals, organized by research category.
info@abcepta.com, and receive a free "I Love Antibodies" mug.

Provided below are standard protocols that you may find useful for product applications.
Background
Members of the CELF/BRUNOL protein family contain two N-terminal RNA recognition motif (RRM) domains, one C-terminal RRM domain, and a divergent segment of 160-230 aa between the second and third RRM domains. Members of this protein family regulate pre-mRNA alternative splicing and may also be involved in mRNA editing, and translation. This gene may play a role in myotonic dystrophy type 1 (DM1) via interactions with the dystrophia myotonica-protein kinase (DMPK) gene. Alternative splicing results in multiple transcript variants encoding different isoforms.
References
Rattenbacher, B., et al. Mol. Cell. Biol. 30(16):3970-3980(2010)
Le Tonqueze, O., et al. Biochem. Biophys. Res. Commun. 394(4):884-889(2010)
Koshelev, M., et al. Hum. Mol. Genet. 19(6):1066-1075(2010)
Tsuda, K., et al. Nucleic Acids Res. 37(15):5151-5166(2009)
Bubenik, J.L., et al. RNA Biol 6(1):73-83(2009)
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.

Ordering Information
Other Products
Shipping Information
