


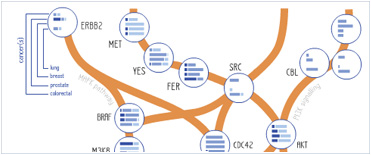







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
UBE2V1 Antibody (C-term)
Purified Rabbit Polyclonal Antibody (Pab)


- SPECIFICATION
- CITATIONS
- PROTOCOLS
- BACKGROUND

Application
| WB, E |
|---|---|
| Primary Accession | Q13404 |
| Other Accession | Q7ZYP0, Q7M767, Q9D2M8, Q15819, Q6PEH5, Q5F3Z3, Q3SZ43, Q9CZY3, Q90879, Q3SZ52, Q9GZW1 |
| Reactivity | Human, Mouse |
| Predicted | Bovine, Chicken, Zebrafish, Rat, Xenopus |
| Host | Rabbit |
| Clonality | Polyclonal |
| Isotype | Rabbit IgG |
| Calculated MW | 16495 Da |
| Antigen Region | 113-145 aa |
| Gene ID | 387522;7335 |
|---|---|
| Other Names | Ubiquitin-conjugating enzyme E2 variant 1, UEV-1, CROC-1, TRAF6-regulated IKK activator 1 beta Uev1A, UBE2V1, CROC1, UBE2V, UEV1 |
| Target/Specificity | This UBE2V1 antibody is generated from rabbits immunized with a KLH conjugated synthetic peptide between 113-145 amino acids from the C-terminal region of human UBE2V1. |
| Dilution | WB~~1:1000 |
| Format | Purified polyclonal antibody supplied in PBS with 0.09% (W/V) sodium azide. This antibody is prepared by Saturated Ammonium Sulfate (SAS) precipitation followed by dialysis against PBS. |
| Storage | Maintain refrigerated at 2-8°C for up to 2 weeks. For long term storage store at -20°C in small aliquots to prevent freeze-thaw cycles. |
| Precautions | UBE2V1 Antibody (C-term) is for research use only and not for use in diagnostic or therapeutic procedures. |
| Name | UBE2V1 |
|---|---|
| Synonyms | CROC1, UBE2V, UEV1 |
| Function | Has no ubiquitin ligase activity on its own. The UBE2V1-UBE2N heterodimer catalyzes the synthesis of non-canonical poly-ubiquitin chains that are linked through Lys-63. This type of poly-ubiquitination activates IKK and does not seem to involve protein degradation by the proteasome. Plays a role in the activation of NF-kappa-B mediated by IL1B, TNF, TRAF6 and TRAF2. Mediates transcriptional activation of target genes. Plays a role in the control of progress through the cell cycle and differentiation. Plays a role in the error-free DNA repair pathway and contributes to the survival of cells after DNA damage. Promotes TRIM5 capsid-specific restriction activity and the UBE2V1- UBE2N heterodimer acts in concert with TRIM5 to generate 'Lys-63'- linked polyubiquitin chains which activate the MAP3K7/TAK1 complex which in turn results in the induction and expression of NF-kappa-B and MAPK-responsive inflammatory genes. Together with RNF135 and UBE2N, catalyzes the viral RNA-dependent 'Lys-63'-linked polyubiquitination of RIGI to activate the downstream signaling pathway that leads to interferon beta production (PubMed:31006531). UBE2V1-UBE2N together with TRAF3IP2 E3 ubiquitin ligase mediate 'Lys-63'-linked polyubiquitination of TRAF6, a component of IL17A-mediated signaling pathway. |
| Cellular Location | Nucleus. Note=Excluded from the nucleolus |
| Tissue Location | Highly expressed in thyroid, pancreas, spinal cord, lymph node, trachea, adrenal gland, bone marrow and pancreas. Detected at low levels in heart, breast, placenta, brain, liver, kidney, stomach and lung. |

Thousands of laboratories across the world have published research that depended on the performance of antibodies from Abcepta to advance their research. Check out links to articles that cite our products in major peer-reviewed journals, organized by research category.
info@abcepta.com, and receive a free "I Love Antibodies" mug.

Provided below are standard protocols that you may find useful for product applications.
Background
The CROC1 isoforms, also known as UBE2V1, show sequence similarity to ubiquitin-conjugating enzymes (UBCs, or E2s) but lack the conserved cysteine residue critical to catalytic activity of E2s.1 Northern blot analysis detected approximately 2.1- and 2.5-kb CROC1 transcripts in all human tissues examined, with the highest levels in brain, skeletal muscle, and kidney. Partial human intestinal epithelial cell cDNAs have been isolated containing the 3-primecoding sequence and 3-prime untranslated region of UBE2V1, also called UEV1.2 UEV1 does not have ubiquitin-conjugating activity in vitro. UEV1 transcripts are downregulated upon differentiation of a colon carcinoma cell line.1 Constitutive expression of exogenous UEV1 protein in these colon carcinoma cells inhibits their capacity to differentiate upon confluence and induces changes in cell cycle behavior associated with inhibition of CDK1. A heterodimeric protein complex has been identified that links TRAF6 to IKK activation.3 Peptide mass fingerprinting analysis revealed that this complex is composed of the ubiquitin conjugating enzyme UBC13 and the UBC-like protein UBE2V1, also called UEV1A. TRAF6, a RING domain protein, functions together with UBC13/UEV1A to catalyze the synthesis of unique polyubiquitin chains linked through lysine-63 (K63) of ubiquitin. Blockade of this polyubiquitin chain synthesis, but not inhibition of the proteasome, prevents the activation of IKK by TRAF6. These results unveil a new regulatory function for ubiquitin, in which IKK is activated through the assembly of K63-linked polyubiquitin chains.
References
Thomson, T.M., et al., Genome Res. 10(11):1743-1756 (2000).
Long, M., Genome Res. 10(11):1655-1657 (2000).
Deng, L., et al., Cell 103(2):351-361 (2000).
Hofmann, R.M., et al., Cell 96(5):645-653 (1999).
Ma, L., et al., Oncogene 17(10):1321-1326 (1998).
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.

Ordering Information
Other Products
Shipping Information
