


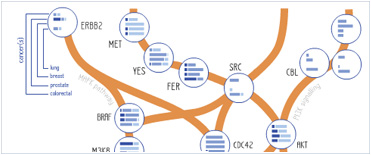







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
HDAC4 Antibody (N-term) Blocking Peptide
Synthetic peptide
- SPECIFICATION
- CITATIONS
- PROTOCOLS
- BACKGROUND
| Primary Accession | P56524 |
|---|---|
| Clone Names | 4013007 |
| Gene ID | 9759 |
|---|---|
| Other Names | Histone deacetylase 4, HD4, HDAC4, KIAA0288 |
| Target/Specificity | The synthetic peptide sequence used to generate the antibody AP1104b was selected from the N-term region of human HDAC4. A 10 to 100 fold molar excess to antibody is recommended. Precise conditions should be optimized for a particular assay. |
| Format | Peptides are lyophilized in a solid powder format. Peptides can be reconstituted in solution using the appropriate buffer as needed. |
| Storage | Maintain refrigerated at 2-8°C for up to 6 months. For long term storage store at -20°C. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Name | HDAC4 (HGNC:14063) |
|---|---|
| Synonyms | KIAA0288 |
| Function | Responsible for the deacetylation of lysine residues on the N-terminal part of the core histones (H2A, H2B, H3 and H4). Histone deacetylation gives a tag for epigenetic repression and plays an important role in transcriptional regulation, cell cycle progression and developmental events. Histone deacetylases act via the formation of large multiprotein complexes. Involved in muscle maturation via its interaction with the myocyte enhancer factors such as MEF2A, MEF2C and MEF2D. Involved in the MTA1-mediated epigenetic regulation of ESR1 expression in breast cancer. Deacetylates HSPA1A and HSPA1B at 'Lys-77' leading to their preferential binding to co-chaperone STUB1 (PubMed:27708256). |
| Cellular Location | Nucleus. Cytoplasm. Note=Shuttles between the nucleus and the cytoplasm. Upon muscle cells differentiation, it accumulates in the nuclei of myotubes, suggesting a positive role of nuclear HDAC4 in muscle differentiation. The export to cytoplasm depends on the interaction with a 14-3-3 chaperone protein and is due to its phosphorylation at Ser-246, Ser-467 and Ser-632 by CaMK4 and SIK1. The nuclear localization probably depends on sumoylation Interaction with SIK3 leads to HDAC4 retention in the cytoplasm (By similarity). {ECO:0000250|UniProtKB:Q6NZM9} |
| Tissue Location | Ubiquitous. |

Thousands of laboratories across the world have published research that depended on the performance of antibodies from Abcepta to advance their research. Check out links to articles that cite our products in major peer-reviewed journals, organized by research category.
info@abcepta.com, and receive a free "I Love Antibodies" mug.

Provided below are standard protocols that you may find useful for product applications.
Background
DNA is wrapped around histone proteins to form nucleosomes and chromatin fiber, a higher-order structure. Chromatin can become alternatively revealed to or concealed from transcription factors. Acetylation of lysine residues induces conformational changes in core histones by destabilizing nucleosomes and allowing transcription factors access to recognition elements in DNA. Deacetylation of histones by histone deacetylases (HDACs) reseals the chromosomal package, leading to a repression of transcription. HDAC4 does not bind DNA directly, but rather through MEF2C and MEF2D. Binding of the N terminus of HDAC4 to MEF2C represses MEF2C transcription activity. The catalytic domain of HDAC4 interacts with HDAC3 via the transcriptional corepressor NCOR2. Experimental conditions leading to the suppression of HDAC4 binding to NCOR2 and to HDAC3 result in loss of enzymatic activity associated with HDAC4, indicating regulation of transcription by bridging the enzymatically active NCOR2-HDAC3 complex and select transcription factors. HDAC4 and MITR contain calmodulin-binding domains that overlap with their MEF2 binding domains. Binding of calmodulin to HDAC4 leads to its dissociation from MEF2, relieving MEF2 from the transcriptional repression by HDAC4. Together, HDAC4, MITR, and CABIN1 constitute a family of calcium-sensitive transcriptional repressors of MEF2. In murine studies, HDAC4, which is expressed in prehypertrophic chondrocytes, interacts with and inhibits the activity of Runx2 in mice, a transcription factor necessary for chondrocyte hypertrophy, establishing HDAC4 as a central regulator of chondrocyte hypertrophy and skeletogenesis.
References
Meinke PT and Liberator P. Curr Med Chem, 8(2): 211- 235 (2001).Nakayama T and Takami Y. J Biochem (Tokyo) 129 (4): 491-499 (2001).Cress, W.D. and Seto, E. J. Cell. Physiol. 184, 1-16 (2000).
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Other Products
Shipping Information
