


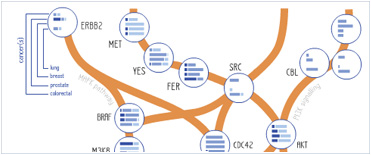







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
Phospho-DNA-PK(S2612) Antibody Blocking peptide
Synthetic peptide
- SPECIFICATION
- CITATIONS
- PROTOCOLS
- BACKGROUND
| Primary Accession | P78527 |
|---|---|
| Clone Names | 5020758 |
| Gene ID | 5591 |
|---|---|
| Other Names | DNA-dependent protein kinase catalytic subunit, DNA-PK catalytic subunit, DNA-PKcs, DNPK1, p460, PRKDC, HYRC, HYRC1 |
| Target/Specificity | The synthetic peptide sequence used to generate the antibody AP3483a was selected from the region of human Phospho-DNA-PK-S2612. A 10 to 100 fold molar excess to antibody is recommended. Precise conditions should be optimized for a particular assay. |
| Format | Peptides are lyophilized in a solid powder format. Peptides can be reconstituted in solution using the appropriate buffer as needed. |
| Storage | Maintain refrigerated at 2-8°C for up to 6 months. For long term storage store at -20°C. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Name | PRKDC |
|---|---|
| Synonyms | HYRC, HYRC1 |
| Function | Serine/threonine-protein kinase that acts as a molecular sensor for DNA damage (PubMed:11955432, PubMed:12649176, PubMed:14734805, PubMed:33854234). Involved in DNA non-homologous end joining (NHEJ) required for double-strand break (DSB) repair and V(D)J recombination (PubMed:11955432, PubMed:12649176, PubMed:14734805, PubMed:33854234, PubMed:34352203). Must be bound to DNA to express its catalytic properties (PubMed:11955432). Promotes processing of hairpin DNA structures in V(D)J recombination by activation of the hairpin endonuclease artemis (DCLRE1C) (PubMed:11955432). Recruited by XRCC5 and XRCC6 to DNA ends and is required to (1) protect and align broken ends of DNA, thereby preventing their degradation, (2) and sequester the DSB for repair by NHEJ (PubMed:15574326, PubMed:11955432, PubMed:12649176, PubMed:14734805, PubMed:33854234). Act as a scaffold protein to aid the localization of DNA repair proteins to the site of damage (PubMed:15574326, PubMed:11955432, PubMed:12649176, PubMed:14734805). The assembly of the DNA-PK complex at DNA ends is also required for the NHEJ ligation step (PubMed:15574326, PubMed:11955432, PubMed:12649176, PubMed:14734805). Found at the ends of chromosomes, suggesting a further role in the maintenance of telomeric stability and the prevention of chromosomal end fusion (By similarity). Also involved in modulation of transcription (PubMed:15574326, PubMed:11955432, PubMed:12649176, PubMed:14734805). As part of the DNA-PK complex, involved in the early steps of ribosome assembly by promoting the processing of precursor rRNA into mature 18S rRNA in the small-subunit processome (PubMed:32103174). Binding to U3 small nucleolar RNA, recruits PRKDC and XRCC5/Ku86 to the small-subunit processome (PubMed:32103174). Recognizes the substrate consensus sequence [ST]-Q (PubMed:15574326, PubMed:11955432, PubMed:12649176, PubMed:14734805). Phosphorylates 'Ser-139' of histone variant H2AX, thereby regulating DNA damage response mechanism (PubMed:14627815, PubMed:16046194). Phosphorylates ASF1A, DCLRE1C, c-Abl/ABL1, histone H1, HSPCA, c-jun/JUN, p53/TP53, PARP1, POU2F1, DHX9, FH, SRF, NHEJ1/XLF, XRCC1, XRCC4, XRCC5, XRCC6, WRN, MYC and RFA2 (PubMed:2507541, PubMed:2247066, PubMed:1597196, PubMed:8407951, PubMed:8464713, PubMed:9362500, PubMed:9139719, PubMed:10026262, PubMed:10467406, PubMed:12509254, PubMed:11889123, PubMed:14612514, PubMed:14599745, PubMed:15177042, PubMed:18644470, PubMed:26666690, PubMed:30247612, PubMed:14704337, PubMed:16397295, PubMed:26237645, PubMed:28712728, PubMed:29478807). Can phosphorylate C1D not only in the presence of linear DNA but also in the presence of supercoiled DNA (PubMed:9679063). Ability to phosphorylate p53/TP53 in the presence of supercoiled DNA is dependent on C1D (PubMed:9363941). Contributes to the determination of the circadian period length by antagonizing phosphorylation of CRY1 'Ser-588' and increasing CRY1 protein stability, most likely through an indirect mechanism (By similarity). Plays a role in the regulation of DNA virus-mediated innate immune response by assembling into the HDP-RNP complex, a complex that serves as a platform for IRF3 phosphorylation and subsequent innate immune response activation through the cGAS-STING pathway (PubMed:28712728). Also regulates the cGAS-STING pathway by catalyzing phosphorylation of CGAS, thereby impairing CGAS oligomerization and activation (PubMed:33273464). Also regulates the cGAS-STING pathway by mediating phosphorylation of PARP1 (PubMed:35460603). |
| Cellular Location | Nucleus. Nucleus, nucleolus |

Thousands of laboratories across the world have published research that depended on the performance of antibodies from Abcepta to advance their research. Check out links to articles that cite our products in major peer-reviewed journals, organized by research category.
info@abcepta.com, and receive a free "I Love Antibodies" mug.

Provided below are standard protocols that you may find useful for product applications.
Background
Protein kinases are enzymes that transfer a phosphate group from a phosphate donor, generally the g phosphate of ATP, onto an acceptor amino acid in a substrate protein. By this basic mechanism, protein kinases mediate most of the signal transduction in eukaryotic cells, regulating cellular metabolism, transcription, cell cycle progression, cytoskeletal rearrangement and cell movement, apoptosis, and differentiation. With more than 500 gene products, the protein kinase family is one of the largest families of proteins in eukaryotes. The family has been classified in 8 major groups based on sequence comparison of their tyrosine (PTK) or serine/threonine (STK) kinase catalytic domains. The STE group (homologs of yeast Sterile 7, 11, 20 kinases) consists of 50 kinases related to the mitogen-activated protein kinase (MAPK) cascade families (Ste7/MAP2K, Ste11/MAP3K, and Ste20/MAP4K). MAP kinase cascades, consisting of a MAPK and one or more upstream regulatory kinases (MAPKKs) have been best characterized in the yeast pheromone response pathway. Pheromones bind to Stem cell surface receptors and activate yeast MAPK pathway.
References
Goudelock, D.M., et al., J. Biol. Chem. 278(32):29940-29947 (2003).Ding, Q., et al., Mol. Cell. Biol. 23(16):5836-5848 (2003).Lucero, H., et al., J. Biol. Chem. 278(24):22136-22143 (2003).Calsou, P., et al., J. Mol. Biol. 326(1):93-103 (2003).Karpova, A.Y., et al., Proc. Natl. Acad. Sci. U.S.A. 99(5):2818-2823 (2002).
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Other Products
Shipping Information
