


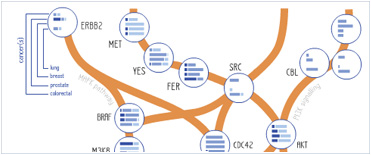







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
Val66Met proBDNF Protein (Mut-human)
A Neurotrophic Factor
- SPECIFICATION
- CITATIONS
- PROTOCOLS
- BACKGROUND
| Storage | -20°C |
|---|---|
| Precautions | Val66Met proBDNF Protein (Mut-human) is for research use only and not for use in diagnostic or therapeutic procedures. |

Thousands of laboratories across the world have published research that depended on the performance of antibodies from Abcepta to advance their research. Check out links to articles that cite our products in major peer-reviewed journals, organized by research category.
info@abcepta.com, and receive a free "I Love Antibodies" mug.

Provided below are standard protocols that you may find useful for product applications.
Background
Brain-derived neurotrophic factor (BDNF) is a neurotrophic factor that bind p75NTR as well as TrkB receptors1,2. BDNF supports the survival of many cell types3-8 and also modulates hippocampal plasticity and hippocampal-dependent memory in cell models and in animals9.
The BDNF gene, like other peptide growth factors, encodes a precursor peptide (proBDNF), which is proteolytically cleaved to form the mature protein10. proBDNF has been shown to be a pro-apoptotic ligand for sympathetic neurons expressing both p75NTR and sortilin11, and to be involved in long-term potentiation (LTP) stage of the memory-related modifications in synaptic transmission12.
A nonconservative single nucleotide polymorphism (SNP) in the human BDNF gene has been identified at nucleotide 196 (G/A) producing an amino acid substitution (Valine to Methionine) at codon 66 (Val66Met, rs 6265). Although located in the 5' pro-BDNF region, this SNP resulted in striking deficits in the cellular distribution and regulated secretion of the mature protein, BDNF and hence in corresponding alterations of human hippocampal function and episodic memory in vivo9.
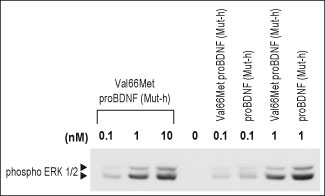
Egan M.F. et al demonstrated the molecular mechanisms that control activity-dependent BDNF secretion and showed that depolarization-dependent secretion of BDNF in hippocampal neurons is significantly impaired when this Val66Met SNP occurs. Using double-staining techniques, they demonstrated that Val-BDNF-containing secretory granules are colocalized with synaptophysin, a marker for synapses. In contrast, Val66Met-BDNF aggregates are accumulated in the cell body and rarely colocalize with synaptophysin. This suggests that even if it can be secreted in small amounts near the cell body through the constitutive pathway, the Met-BDNF protein cannot be secreted at synapses9. Studies of heterozygote BDNF knockout rodents, who presumably have intermediate BDNF levels, demonstrate clear physiological13 and behavioral14 abnormalities, suggesting that secretion levels are critical.
Multiple studies over the recent decades in humans, in vivo in animal models and in vitro found an association between the Val66Met polymorphism and bipolar and unipolar disorders15, Schizophrenia16, 17, anxiety-related behavior18,19and controversial association with ADHD20,21.
The data emerged from the analysis of the Val66Met phenotype in various syndroms and diseases reveal the importance of the pro-region of the BDNF polypeptide, particular Valine66 and perhaps the nearby sequence, in intracellular trafficking and secretion of BDNF.
References
1 . Tolkovsky, A.(1997)Trends Neurosi.20, 1.
2 . Jing, S.et al.(1992)Neuron9, 1067.
3 . Acheson, A.et al.(1995)Nature374,450.
4 . Morse, J.K.et al.(1993)J. Neurosci.13,4146.
5 . Hyman, C.et al.(1991)Nature350,230.
6 . Friedman, B. et al.(1995)J. Neurosci.15,1044.
7 . Meyer, M.et al.(1992)J. Cell Biol.119,45.
8 . Koliatsos, V.E. et al.(1993)Neuron10,359.
9 . Egan, M.F.et al.(2003)Cell112,257.
10 . Seidah, N.G.et al.(1996)FEBS Lett.379,247.
11 . Teng, H.K.et al.(2005)J. Neurosci.25,5455.
12 . Woo, N.H.et al.(2005)Nat. Neurosci.8,1069.
13 . Korte, M.et al.(1995)Proc. Natl. Acad. Sci. U.S.A.92,8856.
14 . Lyons, W.E.et al.(1999)Proc. Natl. Acad. Sci. U.S.A. 96,15239.
15 . Post, R.M.(2007)J. Psychiatric Res.41,979.
16 . Ho, B.C. et al.(2006)Arch. Gen. Psychiatry63,731.
17 . Rosa, A. et al.(2006)Am. J. Med. Gen. B Neuropsych. Gen.141B,135.
18 . Hashimoto, K.(2007)BioEssays29,116.
19 . Chen Z.Y.et al.(2006)Science314,140.
20 . Kent, L.et al.(2005)Mol. Psychiatry10,939.
21 . Sanchez-Mora C.et al.(2010)Am. J. Med. Gen. B Neuropsych. Gen. 153B,512.
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Shipping Information
