


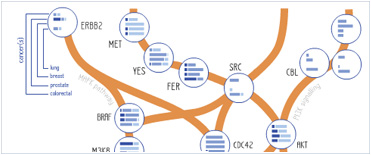







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
Cry1 Antibody (C-term) Blocking Peptide
Synthetic peptide
- SPECIFICATION
- CITATIONS
- PROTOCOLS
- BACKGROUND
| Primary Accession | Q16526 |
|---|---|
| Clone Names | 2080807 |
| Gene ID | 1407 |
|---|---|
| Other Names | Cryptochrome-1, CRY1, PHLL1 |
| Target/Specificity | The synthetic peptide sequence used to generate the antibody AP6134a was selected from the C-term region of human Cry1 . |
| Format | Peptides are lyophilized in a solid powder format. Peptides can be reconstituted in solution using the appropriate buffer as needed. |
| Storage | Maintain refrigerated at 2-8°C for up to 6 months. For long term storage store at -20°C. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Name | CRY1 |
|---|---|
| Synonyms | PHLL1 |
| Function | Transcriptional repressor which forms a core component of the circadian clock. The circadian clock, an internal time-keeping system, regulates various physiological processes through the generation of approximately 24 hour circadian rhythms in gene expression, which are translated into rhythms in metabolism and behavior. It is derived from the Latin roots 'circa' (about) and 'diem' (day) and acts as an important regulator of a wide array of physiological functions including metabolism, sleep, body temperature, blood pressure, endocrine, immune, cardiovascular, and renal function. Consists of two major components: the central clock, residing in the suprachiasmatic nucleus (SCN) of the brain, and the peripheral clocks that are present in nearly every tissue and organ system. Both the central and peripheral clocks can be reset by environmental cues, also known as Zeitgebers (German for 'timegivers'). The predominant Zeitgeber for the central clock is light, which is sensed by retina and signals directly to the SCN. The central clock entrains the peripheral clocks through neuronal and hormonal signals, body temperature and feeding-related cues, aligning all clocks with the external light/dark cycle. Circadian rhythms allow an organism to achieve temporal homeostasis with its environment at the molecular level by regulating gene expression to create a peak of protein expression once every 24 hours to control when a particular physiological process is most active with respect to the solar day. Transcription and translation of core clock components (CLOCK, NPAS2, BMAL1, BMAL2, PER1, PER2, PER3, CRY1 and CRY2) plays a critical role in rhythm generation, whereas delays imposed by post- translational modifications (PTMs) are important for determining the period (tau) of the rhythms (tau refers to the period of a rhythm and is the length, in time, of one complete cycle). A diurnal rhythm is synchronized with the day/night cycle, while the ultradian and infradian rhythms have a period shorter and longer than 24 hours, respectively. Disruptions in the circadian rhythms contribute to the pathology of cardiovascular diseases, cancer, metabolic syndromes and aging. A transcription/translation feedback loop (TTFL) forms the core of the molecular circadian clock mechanism. Transcription factors, CLOCK or NPAS2 and BMAL1 or BMAL2, form the positive limb of the feedback loop, act in the form of a heterodimer and activate the transcription of core clock genes and clock-controlled genes (involved in key metabolic processes), harboring E-box elements (5'-CACGTG-3') within their promoters. The core clock genes: PER1/2/3 and CRY1/2 which are transcriptional repressors form the negative limb of the feedback loop and interact with the CLOCK|NPAS2-BMAL1|BMAL2 heterodimer inhibiting its activity and thereby negatively regulating their own expression. This heterodimer also activates nuclear receptors NR1D1/2 and RORA/B/G, which form a second feedback loop and which activate and repress BMAL1 transcription, respectively. CRY1 and CRY2 have redundant functions but also differential and selective contributions at least in defining the pace of the SCN circadian clock and its circadian transcriptional outputs. More potent transcriptional repressor in cerebellum and liver than CRY2, though more effective in lengthening the period of the SCN oscillator. On its side, CRY2 seems to play a critical role in tuning SCN circadian period by opposing the action of CRY1. With CRY2, is dispensable for circadian rhythm generation but necessary for the development of intercellular networks for rhythm synchrony. Capable of translocating circadian clock core proteins such as PER proteins to the nucleus. Interacts with CLOCK-BMAL1 independently of PER proteins and is found at CLOCK-BMAL1-bound sites, suggesting that CRY may act as a molecular gatekeeper to maintain CLOCK-BMAL1 in a poised and repressed state until the proper time for transcriptional activation. Represses the CLOCK-BMAL1 induced transcription of BHLHE40/DEC1. Represses the CLOCK-BMAL1 induced transcription of ATF4, MTA1, KLF10 and NAMPT (By similarity). May repress circadian target genes expression in collaboration with HDAC1 and HDAC2 through histone deacetylation. Mediates the clock-control activation of ATR and modulates ATR-mediated DNA damage checkpoint. In liver, mediates circadian regulation of cAMP signaling and gluconeogenesis by binding to membrane-coupled G proteins and blocking glucagon-mediated increases in intracellular cAMP concentrations and CREB1 phosphorylation. Inhibits hepatic gluconeogenesis by decreasing nuclear FOXO1 levels that down-regulates gluconeogenic gene expression (By similarity). Besides its role in the maintenance of the circadian clock, is also involved in the regulation of other processes. Represses glucocorticoid receptor NR3C1/GR-induced transcriptional activity by binding to glucocorticoid response elements (GREs). Plays a key role in glucose and lipid metabolism modulation, in part, through the transcriptional regulation of genes involved in these pathways, such as LEP or ACSL4 (By similarity). Represses PPARD and its target genes in the skeletal muscle and limits exercise capacity (By similarity). Plays an essential role in the generation of circadian rhythms in the retina (By similarity). Represses the transcriptional activity of NR1I2 (By similarity). |
| Cellular Location | Cytoplasm. Nucleus Note=Translocated to the nucleus through interaction with other clock proteins such as PER2 or BMAL1. {ECO:0000250|UniProtKB:P97784} |

Thousands of laboratories across the world have published research that depended on the performance of antibodies from Abcepta to advance their research. Check out links to articles that cite our products in major peer-reviewed journals, organized by research category.
info@abcepta.com, and receive a free "I Love Antibodies" mug.

Provided below are standard protocols that you may find useful for product applications.
Background
Various biochemical, physiological and behavioural processes display circadian rhythms controlled by an internal biological clock. The central ??ears??driving this clock appear to be composed of an autoregulatory transcription/posttranslation-based feedback loop. Cryptochrome 1 (CRY1) and 2 (CRY2) are DNA-binding flavoproteins that bear some homology to blue-light receptors and photolyases. In Drosophila, CRY is a photoreceptor for the circadian clock where it binds to the clock component TIM in a light-dependent fashion and blocks its function. Mammalian CRY1 and CRY2 function via light-independent interactions with circadian genes CLOCK and BMAL1, as well as with PER1, PER2, and TIM. They seem to act as light-independent components of the circadian clock and likely regulate Per1 transcriptional cycling via interactions with both the activator and its feedback inhibitors. Mutant mice not expressing the Cry1 or Cry2 protein display accelerated and delayed periodicity of locomotor activity, respectively. It appears that the combination of both proteins working together is essential to synchronize the organism to circadian phases. A critical balance between Cry1 and Cry2 is required for proper clock function; in complete darkness, double-mutant mice present with instantaneous arrhythmicity, indicating the absence of an internal circadian clock.
References
Griffin, E.A. Jr., et al., Science 286(5440):768-771 (1999).van der Horst, G.T., et al., Nature 398(6728):627-630 (1999).Kobayashi, K., et al., Nucleic Acids Res. 26(22):5086-5092 (1998).Hsu, D.S., et al., Biochemistry 35(44):13871-13877 (1996).van der Spek, P.J., et al., Genomics 37(2):177-182 (1996).
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Other Products
Shipping Information
